Research Study 1

Fig. 1. Ventral view of abdominal pleopods of Ligia pallasii

Fig. 2. Comparison of gas-exchanging organs in selected oniscid isopods Ligia pallasii, Oniscus asellus, Porcellio scaber, and Armadillidium vulgare
The branchial chamber in an isopod consists of five pairs of pleopods1. In marine forms the pleopods are double, with hemolymph-filled sacs that act as simple "diffusion gills" and outer flattened parts that are protective. In terrestrial forms, including the semiterrestrial Ligia pallasii, each pleopod is also double, with an innermost endopod and an outermost protective exopod. The exopods fit snugly together to form a protective cover for the branchial chamber (Fig. 1). In all terrestrial genera, the endopods are similarly structured, fleshy, hemolymph-filled sacs, while the exopods, in addition to fulfilling a protective role, are variously modified for gas exchange. Note in Fig. 22 an apparent evolutionary3 transition from sac-like structures in Ligia pallasii, to corrugated lung-like cavities in Oniscus asellus, to a gradation of air-filled invaginations called “pseudotracheae” in Porcellio scaber and Armadillidium vulgare. This increasing specialisation is reflected in an evolution to more terrestrial habitats occupied, from splash-zone marine habitats for ligiids, to wet-forest habitats for oniscids, and to more arid habitats for porcellionids and armadillidids.
NOTE1 lit. “swim legs” G., referring to the fact the these abdominal appendages are used for swimming in various crustaceans, including aquatic isopods
NOTE2 in addition to the evolutionary progression they represent, the three terrestrial species shown here are selected because, along with L. pallasii, they can readily be collected within a few hundred meters of one another where human habitations are found close to rocky shorelines
NOTE3 although some authors believe that the different modes of gas exchange in oniscids have evolved independently, it is convenient for explanatory purposes to consider them as an evolutionary transition as shown here
Carefoot & Taylor 1995 In, Terrestrial Isopod Biology (Alikhan, Ed.) AA Balkema, Rotterdam
Research Study 2
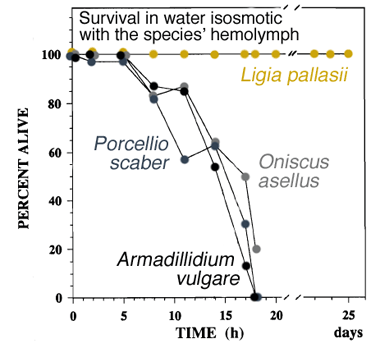
Fig. 1. Survival of oniscid isopods during immersion
While Ligia pallasii tend not to spend time in seawater, there may be occasions when they enter this environment. For example, when foraging for seaweeds or during exposure to storm surge in their supralittoral habitat, they may on occasion be washed by waves into the sea. Ligia may also enter shallow tidepools voluntarily or, more precipitously, to tumble or jump from rocks and cliff edges when in escape mode. Experiments at Bamfield Marine Sciences Centre, British Columbia show that L. pallasii is able to survive indefinitely in seawater, with oxygen consumption being maintained at a high level (Fig. 1). More specialised terrestrial forms such as the pillbug Armadillidium vulgare and other common woodlice species (Porcellio scaber and Oniscus asellus) are incapable of long-duration aquatic gas exchange and die in less than a day when immersed.
NOTE tests are done at 15oC in 100% seawater for L. pallasii and, for the other oniscid species, in seawater diluted to match the species' hemolymph osmolality (varies from about 60 - 65% seawater). This latter minimises or avoids additional osmotic stresses
Carefoot & Taylor 1995 In, Terrestrial Isopod Biology (Alikhan, ed.) AA Balkema, Rotterdam
Research Study 3

Fig. 1. Comparison of oxygen uptake in water in selected oniscid isopods Ligia pallasii, Oniscus asellus, Porcellio scaber, and Armadillidium vulgare
A comparison at the University of British Columbia of aquatic oxygen uptake of the semiterrestrial Ligia pallasii and the three common terrestrial species featured above is also revealing, not just for reason that the level of underwater gas-exchange proficiency tends to reflect the degree of terrestriality, but also for the fact that there is at least some retention in terrestrial isopods of an ancestral ability for underwater oxygen uptake (Fig. 1). Moreover, all species can survive several hours of immersion in fresh water, sufficient to withstand temporary rain deluge. Ligia pallasii’s superior capabilities to survive and respire in seawater reflect its greater similarity to aquatic ancestors, and an evolutionary series showing gas-exchange capability and ability to survive in water for four common west-coast oniscid species would be as follows: L. pallasii > O. asellus = P. scaber = A. vulgare.
NOTE the authors use seawater isosmotic with the internal body fluids of each species to reduce possible confounding effects of osmotic stress
Taylor & Carefoot 1993 Can J Zool 71: 1378
Research Study 4

Figs. 1 - 3. Comparison of metabolic conformation/regulation in three oniscid species. Yellow lines indicate metabolic conformation, where oxygen utilisation is in direct proportion to oxygen content in the air. Any useage above this line, such as by Armadillidium vulgare, indicates an ability to regulate uptake
How does one assess the efficacy of different gas-exchange mechanisms in the evolutionary array of terrestrial isopods shown above? This question is answered by researchers at Claremont Colleges, California who compare oxygen consumption in several species at different levels of oxygen concentration (PO2) and humidity. Results for the first are clear-cut, with Ligia occidentalis acting as a metabolic conformer, Alloniscus perconvexus possibly a partial regulator, and Armadillidium vulgare a strong regulator except at the lowest PO2 level tested (see Figs. 1 - 3, where normal air is 21% O2). The species A. vulgare has a highly evolved gas-exchange system and is arguably the most well-adapted of the terrestrial species listed here. Humidity effects on oxygen uptake are less clear, except for Ligia and Oniscus asellus after being subjected to 10% dehydration, where oxygen uptake significantly decreases (data not shown). In comparison, dehydration effects in Armadillidium are not significant. The researchers show in other experiments that Ligia, a conformer, does not accumulate lactate under mild hypoxia indicating that anaerobic metabolism is not occurring. In contrast Alloniscus perconvexus a partial regulator, exhibits high lactate accumulation in hypoxic conditions. This species lives intertidally in sand burrows, and the authors conjecture that anaerobiosis might be an adaptation to periodic hypoxic exposure in the burrows and the cost of burrowing itself. In comparison, the metabolic regulating Armadillidium vulgare shows no significant anaerobic adaptation to hypoxia. Note that this highly adapted terrestrial species can regulate down to 10% oxygen content. How is this of advantage? Although not mentioned by the authors, Armadillidium rolls up into a ball when disturbed, and it may be that in this position oxygen becomes temporarily limiting owing to occlusion of the gas-exchanging pseudotracheal openings.
NOTE of six species included in the study, only Ligia occidentalis (hemolymph-filled pouches), Alloniscus perconvexus (thinning of the gas-exchange cuticle), Oniscus asellus (similarly thinned cuticle), and Armadillidium vulgare (pseudotracheae or air-filled tubules) are included in this summary, representing three distinctly different morphologies of gas-exchange surfaces
NOTE measured at Relative Humidities of 90 and 55% with a third treatment involving dehydration to 10% less body mass, followed by measurement of oxygen uptake in 55% humidity
Wright & Ting 2006 Comp Biochem Physiol A 145: 235
 Isopods
Isopods 