 Sea stars
Sea stars 
Locomotion by tube-foot crawling in echinoderms involves the water-vascular system that in sea stars originates from a central ring-canal hub and extends distally to the arm tips. The tube feet connect to the water-vascular system via lateral canals, radial canals, and ring canal (Fig. 1). From the ring canal a calcified “stone” canal (not labeled in the drawing) leads to the madreporite. The madreporite is visible externally as a porous plate, and is often distinctively coloured. Because of the physical nature of the connection of the madreporite with the water-vascular system, it has long been thought to act as a conduit for entry of seawater for operation of the tube feet and/or for body-volume regulation. However, unequivocal evidence for this is known only from relatively recent
Research Study 1
Studies on madreporite function in the ochre star Pisaster ochraceus (Fig. 1) show that individuals with
Ferguson 1992 Biol Bull 183: 482
Ferguson 1996 Biol Bull 191: 431
Research Study 2


After bathing Leptasterias hexactis in seawater containing fluorescent microbeads (0.2µm dia) for 48h, high concentrations of beads are found in the lumen and lining of the tube feet, the ampullae, and within the Tiedemann’s bodies. The route taken is from madreporite (Figs. 1 - 2) to stone canal, then radially outwards to ampullae and tube feet. The flow rate is small (2 - 3 µl per g animal per h) but, as noted in Research Study 1 above, is considered by the authors necessary for sustaining the volume of the perivisceral coelomic fluid. Detailed studies on the fine structure of madreporites and stone canals in sea stars suggest that the madreporite is not just a simple sieve-like entrance to the water-vascular system; rather, it has abundant secretory cells, phagocytic coelomocytes, and an extensive complex of nerves. The authors conjecture that the madreporite may utilise the strong ciliary pumping of the stone canal to cleanse coelomic fluids drawn in from the
Ferguson & Walker 1991 J Morphol 210: 1
Research Study 3
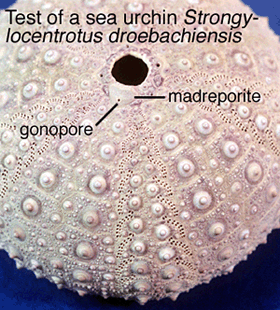

Just for comparison, the madreporite of sea urchins Strongylocentrotus droebachiensis consists of 300 - 400 pore canals lined with cilia (Fig. 1). However, the pores are so small that, in total, they represent only about 0.1mm2. Studies at Friday Harbor Laboratories, Washington using fluorescent microbeads (0.2µm dia) placed for 5d around these openings show inward movement of fluid at an estimated rate of 0.1ml per day for a 200g animal. Most of the beads are caught up within the stone canal (Fig. 2), but a few make it as far as the tube feet. Is this enough of a flow rate to be functionally significant? By blocking the madreporite with cement, the author is able to show a significant decrease in mass over 27d and credits this to less food being present in the gut. While diminished feeding could result from gradual loss of tube-feet activity through decrease in volume of the water-vascular system, as suggested by the author, could it also possibly be an effect of scraping and plugging treatment itself, especially if the madreporite functions in ways other than water-volume regulation.